摘要:
关键词:肠杆菌肽;抗菌;抗炎;肠道健康
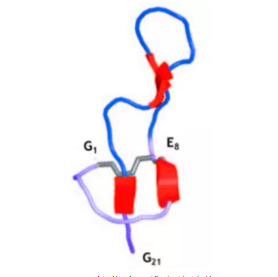
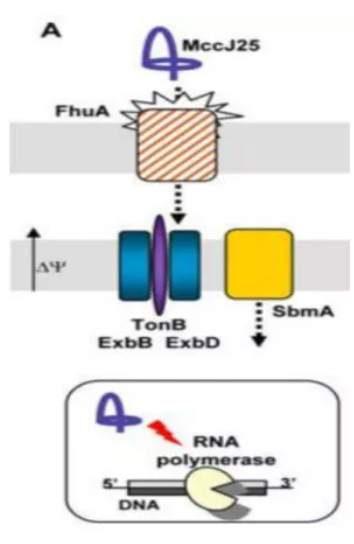
抗生素在养殖业中的使用非常普遍,但滥用抗生素造成的药物残留和细菌耐药性问题已经严重威胁到人体健康和环境的发展,美国每年有200万人遭受抗生素耐药性感染,且其中有2.3万人由于抗生素耐药性感染而死亡[1]。目前很多国家都出台了禁用抗生素的政策,抗生素替代问题也引起了人们的重视。自1981年,瑞典科学家Boman 等从惜古比天蚕蛹中首次分离出天蚕素以来,人们在动植物及细菌、真菌体内发现了超过3400种抗菌肽。抗菌肽不仅来源广泛,而且热稳定性高、抑菌谱广、耐酸碱和蛋白酶,能够有选择性地杀灭细菌或真菌。此外,还具有抗病毒、抗氧化、促生长、调节免疫等多种生物学功能。 肠杆菌肽(Lasso Peptides)由美国普林斯顿大学于1993年从肠道细菌中分离得到的一种新型抗菌肽,由21个氨基酸组成,具有套索状结构,无溶血性,结构稳定,对胃蛋白酶、胰蛋白酶以及高温环境有着良好耐受性,能够有效的杀灭大肠杆菌、沙门氏菌、志贺氏菌等革兰氏阴性菌,且不易产生耐药性[2,3]。目前,已经有大量动物临床试验证明,肠杆菌肽在替代抗生素添加到畜禽饲料中具有较好的抗菌消炎、增强肠道屏障、改善肠道菌群的效果。相信肠杆菌肽未来在畜牧养殖领域会发挥越来越重要的作用,减少动物性食品的药物残留,为人类健康保驾护航。 肠杆菌肽国内外研究进展 肠杆菌肽的理化性质 图1 肠杆菌肽三维立体结构 抗菌肽的抑菌机制 肠杆菌肽的摄取机制:外膜蛋白FhuA的表达是成熟肠杆菌肽进入细胞的前提和所必需的蛋白。肠杆菌肽首先与FhuA受体的外环结合,然后内膜TonB-ExbB-ExbD复合物和SbmA将其转运到细菌细胞质中[4-6],进而发挥细胞内抗菌机制。肠杆菌肽主要以Salmnella和E coli 为靶点,作用机制是:一,通过抑制RNA聚合酶活性来干扰细菌mRNA以及蛋白质的合成。RNA 聚合酶的β亚基发生突变后,其对肠杆菌肽产生耐受性,从而确定其靶位点为RNA 聚合酶[7];肠杆菌肽与RNA聚合酶次级通道的结合会阻碍核苷酸进入酶的活性位点,从而抑制 mRNA的形成和进一步蛋白质的合成[8]。二,肠杆菌肽通过改变细胞膜的通透性,使细菌细胞膜破碎,内容物外渗,导致病原菌死亡。如图2所示。 图2 肠杆菌肽的摄取及抑菌机制 肠杆菌肽具有广谱的抗革兰氏阴性菌活性,包括致病性大肠杆菌、沙门氏菌和志贺氏菌。据报道,肠杆菌肽对沙门氏菌的MIC为3.12 μg/mL,对大肠杆菌O157:H7的MIC为1 μg/mL,0.3%的肠杆菌肽对大肠杆菌 ATCC25922的MIC为300μg/mL,对大肠杆菌AZ1、大肠杆菌W1和W2的MIC均为200μg/mL,对沙门氏菌SK226和沙门氏菌1791的MIC均为100μg /mL[9-11]。而且肠杆菌肽对大肠杆菌和沙门氏菌野毒株也具有明显的杀灭作用,随着肠杆菌肽浓度的变大,其抑菌圈直径不断变大。Yu等[12-13]研究发现肠杆菌肽不仅对大肠杆菌和沙门氏菌具有显著的抑杀效果,而且适量的肠杆菌肽还可以增加乳酸杆菌和双歧杆菌等有益数量,并且还对肠杆菌肽对大肠杆菌和沙门氏菌的最小抑菌浓度做了详细的比较,如表1、表2。 表1 肠杆菌肽对标准致病菌的最小抑菌浓度

注:aLKFZ菌株由北京生物饲料添加剂重点实验室提供。临床来源菌株SWI、BRO和CATT由中国农业大学兽医学院兽药检测中心提供。SWI、BRO和CATT临床来源的菌株是耐抗生素(林可霉素和替米考星)的沙门氏菌和大肠杆菌。b所有MIC和MBC值代表3个独立的试验均值。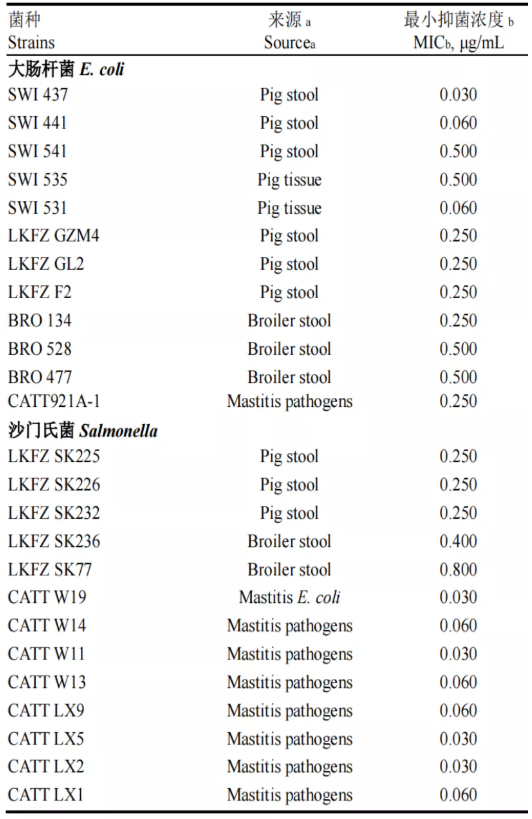
肠杆菌肽除了抗菌消炎的作用,还具有提高采食量、降低料肉比、提高生产性能和调节免疫的功能。有研究发现,在饲料中添加500g/t 肠杆菌肽可以提高断奶仔猪的日采食量、日增重、降低料肉比、降低腹泻率、使仔猪毛色变优[17-19]。孙何军等[20]研究报道,在基础饲粮中添加100 g/t的肠杆菌肽可以使蛋鸡产蛋率提高3.88%、脏蛋率降低12.68%、料蛋比下降0.07、死淘率降低40%、破蛋率降低14.63%,添加肠杆菌肽能显著提高蛋鸡的产蛋性能。关静姝等[21]研究发现,在肉鸡基础日粮中添加300g/t 肠杆菌肽增加了肉鸡的出栏体重和平均日增重,降低了料重比,提高了饲料的转化率。卜艳玲等[22]通过在仔猪饲喂的基础饲粮中分别添加30 g/t硫酸黏菌素、300 g/t天蚕素抗菌肽以及300、400和500 g/t肠杆菌肽对比发现,添加500 g/t 肠杆菌肽对提高断奶仔猪生产性能、免疫性能和抗氧化能力均有显著效果,且优于硫酸黏菌素。程振峰[23]研究发现肠杆菌肽(500g/t)、微生态制剂(500g/t)+肠杆菌肽(500g/t)组合对提高生长育肥猪平均日增重、降低料重比、提高仔猪血液中白细胞总数和淋巴细胞比率方面也具有显著效果。
肠杆菌肽的稳定性与安全性
肠杆菌肽是利用蛋白工程技术在植物乳杆菌中高效生产的一种抗菌肽,肠杆菌肽的结构包含一个由Gly1的ɑ氨基和Glu8的γ羧基之间形成的内酰胺键连接的8个氨基酸残基组成的(Gly1-Glu8)套索结构环,该多肽的尾部(Tyr9-Gly21)从这个套索环中穿过,且Ple19和Tyr20分别卡于环的两侧,在空间上通过非共价作用将这个尾部固定在环内,这种独特的结构使其性质非常稳定。刘扬科等[24]通过体外进行的肠杆菌肽对大肠杆菌和沙门氏菌的抑菌效果研究发现,在人工胃液、肠液和热处理的环境下,0.3%肠杆菌肽对肠杆菌和沙门氏菌的MIC为128 μg/mL,表明肠杆菌肽具有很强的稳定性。卜艳玲[22]也发现了类似的结果。表明肠杆菌肽对高温、酸碱、蛋白酶等具有很好的耐受性。 目前,还未见有使用肠杆菌肽后出现不良反应的报道。Lopez等[9]研究报道,肠杆菌肽体外经过24 h在全血、血浆以及血清中仍保持完全活性,且不会出现溶血现象,能显著抑制全血、同源的血浆和血清中沙门氏菌的数量。肠杆菌肽与传统的抗生素抗菌作用机理具有很大的不同点,不易产生耐药性,作为新药开发来对抗细菌感染的潜力巨大[25,26]。
肠杆菌肽应用研究前景分析
目前,肠杆菌肽的研发和应用已经日趋成熟,一些抗菌肽产品已经规模化生产并投入到了生产一线,且得到了较好的反馈效果。除了肠杆菌肽原有的一些生物功能以外,通过生物工程等先进技术,对肠杆菌肽进行改造,发挥肠杆菌肽更优质的疗效和特异性功能以后可能会成为一个发展趋势。提高肠杆菌肽的特异性可以通过减少或改变胃肠道细菌群落中的微生物群的组成,有效的预防原发性和机会性感染[27]。肠杆菌肽未来在畜牧业、化妆品和食品防腐等领域的应用前景将会更加的广泛。
[1] Martens E, Demain AL. 2017 The antibiotic resistance crisis, with a focus on the United States[J]. Antibiot,70:520-526. [2] Bayro, Marvin J, Mukhopadhyay J, Swapana G V, et al. Structure of antibacterial peptide microcin J25: A 21-Residue lariat protoknot[J]. J Am Chem Soc, 2003, 125(41): 12382-12383. [3] Maria R R, Beatriz F A, Raúl A S, et al. The antibacterial action of microcin J25: evidence for disruption of cytoplasmic membrane energization in Salmonella newport[J]. FEMS Microbiol Lett, 2001, 204(2): 265-270. [4] Rosengren K J, Clark R J, Daly N L, Go ansson U, Jones A, Craik DJ Microcin J25 has a threaded sidchain-no-backbone ring srcture and not a ha-o-tail cyclized backbon[J]. Jourmal oftheAmerican Chemical Society, 2003, 125(41):12464-12474. [5] Destoumieux-Garzon D, Duquesne S, Peduzzi J, Goulard C, Desmadril M, Letellier L,Rebufat S, Boulanger P. The iron-siderophore transporter FhuA is the receptor for the antimicrobialpeptide microcin J25: Role of the microcin Valn-Pro16 B-hairpin region in the recognition mechanism[J].The Biochemical Journal, 2005, 389(3):869-876. [6] Salomon R A, Farias R N. The pepide antibiotic microcin J25 is imported through the TonBpathway and the SbmA protein[J]. Jourmal of Bacteriology, 1995, 7711.3323-3325. [7] Yuzenkova J, Delgado M, Nechaev S, et al. Mutations of bacterial RNA polymerase leading to resistance to microcin j25[J]. J Biol Chem, 2002, 277(52): 50867-50875. [8] Mukhopadhyay J, Sineva E, Knight J , et al. Antibacterial peptide microcin J25 inhibits transcription by binding within and obstructing the RNA polymerase secondary channel[J]. Mol Cell, 2004, 14(6): 739-752. [9] Lopez F E, Vincent P A, Zenoff A M, et al. Efficacy of microcin J25 in biomatrices and in a mouse model of Salmonella infection[J]. J Antimicrob Chemother, 2007, 59(4): 676-680. [10]熊海涛,刘扬科,张志华,等.无抗日粮中添加肠杆菌肽对仔猪生长性能和粪便微生物的影响[J].中国畜牧杂志,2019,55(02):118-121. [11] Sable S, Pons A M, Gendron-Gaillard S, et al. Antibacterial activity evaluation of microcin J25 against diarrheagenic Escherichia coli[J]. Appl Environ Microbiol, 2000, 66(10): 4595-4597. [12]H.T. Yu, X. L. Ding, N. Li,et al.Dietary supplemented antimicrobial peptide microcin J25 improves the growth performance, apparent total tract digestibility, fecal microbiota, and intestinal barrier function of weaned pigs[J]. J. Anim. Sci. 2017.95:5064–5076. [13]于海涛. 抗菌肽MicrocinJ25抗肠毒素大肠杆菌感染的作用及其机制研究[D]. 北京:中国农业大学,2020. [14] Gang Wang, Qinglong Song, Shuo Huang, et al. Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation,and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella[J].Animals,2020 Feb 21;10(2):345. [15] Zasloff M. Antimicrobial peptides of multicellular organisms[J]. Nature, 2013, 415(6870): 389-395. [16] V Portrait, S Gendron-Gaillard, G Cottenceau,et al.Inhibition of pathogenic Salmonella enteritidis growth mediated by Escherichia coli microcin J25 producing strains[J].Can J Microbiol. 1999 Dec;45(12):988-94. [17]赵青海, 胡毅. 肠杆菌肽替代土霉素钙对断奶仔猪生长性能的影响[J]. 湖南饲料,2019,06:30-32. [18]关静姝,李成功,等. 日粮中添加肠杆菌肽对断奶仔猪生产性能的影响[J]. 广东饲料,2016,25(5):21-23. [19]李希,何涛,等. 日粮中添加肠杆菌肽对保育猪生产性能的影响[J]. 饲料博览, 2016(2):1-4. [20]孙何军,刘成宏,关静姝,等. 日粮中添加肠杆菌肽对蛋鸡产蛋性能的影响[J]. 饲料工业,2016,37(13):28-30. [21] 关静姝,刘成宏,等. 日粮中添加肠杆菌肽对肉鸡生长性能的影响[J]. 饲料与畜牧,2015,(12):50-51. [22]卜艳玲,陈静,李建涛,等.饲粮中添加肠杆菌肽对断奶仔猪生产性能和血清生化指标的影响[J].动物营养学报,2018,30(2):696-706. [23]程振峰.不同抗生素替代物对生长肥育猪生长、肠道形态与发育影响的研究[D].山东泰安:山东农业大学,2018. [24]关静姝,赵裕川,刘扬科,等. 肠杆菌肽对大肠杆菌和沙门氏菌的体外抑菌效果研究[J]. 中国畜牧杂志,2018,54(5):122-125. [25] Rania S, Sahar A, Kamaljit K. NGR Peptide ligands for targeting CD13/APN identified through peptide array screening resemble fibronectin sequences[J]. ACS Comb Sci, 2012, 14(11): 590-599. [26] Mathavan I, Zirah S, Mehmood S, et al. Structural basis for hijacking siderophore receptors by antimicrobial lasso peptides[J]. Nat Chem Biol, 2014, 10(5): 340-342. [27] Guinane C M, Cotter P D. Role of the gut microbiota in health and chronic gastrointestinal disease: understanding a hidden metabolic organ[J]. Therap. Adv. Gastroenterol. 2013, 6:295–308. 撰稿人:宋志恒






